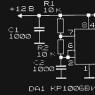
Апоптоз и пути его реализации. Апоптоз
Федеральное государственное общеобразовательное учреждение высшего профессионального образования
МОСКОВСКАЯ ГОСУДАРСТВЕННАЯ АКАДЕМИЯ ВЕТЕРИНАРНОЙ МЕДИЦИНЫ И БИОТЕХНОЛОГИИ им К.И. СКРЯБИНА
ПРОВЕРИЛ
Преподаватель иммунологии
_________________________
«__» ________ 2007 г.
РЕФЕРАТ ПО ИММУНОЛОГИИ
на тему:
АПОПТОЗ И ПУТИ ЕГО РЕАЛИЗАЦИИ
Исполнитель: _______Н.Н. Шитов
Москва
АПОПТОЗ: ВВЕДЕНИЕ.................................................................................................. 4 стр
АПОПТОЗ: ВОЗМОЖНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ ЭТАПОВ.......................... 5 стр
На клетку действуют сигналы, индуцирующие апоптоз ....................................5 стр
Сигналы апоптоза передается внутрь клеток .................................................6 стр
Сигналы апоптоза меняют работу генов .........................................................6 стр
Клетка анализирует сигналы ............................................................................6 стр
П рокаспазы: активация .........................................................................................7 стр
Последовательность действия про- и антиапоптозных сигналов ......................8 стр
Апоптоз: фаза экзекуции (внеядерная) ................................................................9 стр
Введение .............................................................................................................9 стр
Ф аза высвобождения .........................................................................................9 стр
В ведение .......................................................................................................9 стр
Ключевые белки .........................................................................................10 стр
Белки фокальной адгезии........................................................................10 стр
Актиновые регуляторы ............................................................................10 стр
Микротрубочки (mt) ................................................................................10 стр
Эффекторы ………………………………………………………………..10 стр
протеазы ………………………………………………………………...10 стр
калпаины ………………………………………………………………..10 стр
киназы ………………………………………………………………….10 стр
Фаза Блеббинга ………………………………………………………………11 стр
В ведение ………………………………………………………………….11 стр
К лючевые цитоскелетные белки ………………………………………..11 стр
миозины …………………………………………………………………11 стр
актин ……………………………………………………………………..11 стр
актин-связывающие белки ……………………………………………..11 стр
эффекторы ………………………………………………………………11 стр
ATP ……………………………………………………………………… 11 стр
Апоптоз: фаза экзекуции: фаза конденсации ……………………………… 12 стр
Введение ………………………………………………………………….12 стр
К лючевые белки ………………………………………………………….12 стр
Э ффекторы ………………………………………………………………..12 стр
протеазы ………………………………………………………………...12 стр
PAK киназы ……………………………………………………………...12 стр
Т рансглютаминаза ………………………………………………………12 стр
Апоптоз: роль внеядерной фазы экзекуции ………………………………...12 стр
АПОПТОЗ: ПУТИ РЕАЛИЗАЦИИ…………………………………………………...13 стр
Апоптоз, опосредованный рецепторами ……………………………………… 13 стр
В ведение ……………………………………………………………………….13 стр
Р ецепторы апоптозные ……………………………………………………….13 стр
Р ецепторы смерти …………………………………………………………13 стр
рецепторы смерти: общие сведения ……………………………………13 стр
CD95 рецептор: передача сигнала ……………………………………...14 стр
FADD и индукция апоптоза через CD 95 ...............................................15 стр
TNFR1 : передача сигнала .........................................................................15 стр
DR3 : передача сигнала ..............................................................................16 стр
DR3 и DR5: модуляция передачи сигнала apo2l ложными рецепторами ...............................................................................................17 стр
DCR рецепторы DCR1 и DCR2 ................................................................17 стр
апоптоз, индуцированный FAS рецептором ...........................................18 стр
Лиганды рецепторов смерти ............................................................................19 стр
Р ецепторы смерти для индукции апоптоза в опухолях .................................19 стр
А поптоз: митохондриальный механизм реализации .........................................19 стр
В ведение .............................................................................................................19 стр
Происхождение и апоптоз ................................................................................19 стр
М итохондрии и апоптоз, незавмсимый от каспаз ..........................................20 стр
З апуск смерти клеток ........................................................................................21 стр
Н арушение электронного транспорта ......................................................21 стр
В ысвобождение цитохрома С...................................................................21 стр
роль высвобождения цитохрома c в апоптозе .......................................21 стр
последствия высвобождения цитохрома С .......................................... 22 стр
цитохром c , роль в апоптозе, эволюционный аспект ........................... 22 стр
нарушение окислительного потенциала и образование пероксидов .... 23 стр
Митохондрии: нарушение работы мембран при апоптозе ..........................23 стр
PT поры во внутренней мембране, разрушение внешней мембраны и апоптоз ........................................................................................................... 24 стр
Разрушение внешней мембраны из-за гиперполяризации.....................25 стр
Открытие большого канала во внешней мембране..................................25 стр
Митохондрии и белки BCL-2 при апоптозе ...................................................25 стр
В лияние белков BCL-2 ...............................................................................25 стр
Подобие каналообразующим белкам ........................................................26 стр
М еханизм действия белков семейства BCL-2 .........................................26 стр
Б елки BCL-2, подобие каналообразующим белкам, эволюционный аспект ............................................................................................................. 26 стр
AIF белок: митохондриальный эффектор апоптоза независимый от каспаз ............................................................................................................. 27 стр
А поптоз: другие механизмы реализации ............................................................27 стр
Апоптоз с участием рецепторного и митохонриального механизмов .........28 стр
А поптоз с участием эндоплазматического ретикулума ................................28 стр
А поптоз, вызванный цитотоксическими лимфоцитами ...............................28 стр
Апоптоз, вызванный нарушением адгезии клеток........................................28 стр
А поптоз эритроцитов ........................................................................................28 стр
А поптоз: маркеры ..................................................................................................28 стр
ИСПОЛЬЗУЕМАЯ ЛИТЕРАТУРА...............................................................................30 стр
АПОПТОЗ: ВВЕДЕНИЕ
Термин"апоптоз", предложенный в 1972 г. английскими учеными J.F.R. Кеrr, А.Н. Wyllie и A.R. Currie, состоит из двух греческих слов и означает в буквальном смысле "отделение лепестков от цветов", а применимо к клетке - особый тип смерти путем разделения ее на части (" апоптозные тельца
"), которые впоследствии фагоцитируются соседними клетками разного типа.
Термин "программированная клеточная смерть" отражает функциональное назначение этого процесса, представляющего естественную часть жизни многоклеточного организма, связанного с метаморфозом и развитием.
В генетическом аппарате многоклеточных организмов - животных, растений и грибов заложена программа гибели клеток. Это специальная программа, которая при определенных обстоятельствах может привести клетку к гибели. При нормальном развитии эта программа направлена на удаление избыточно образовавшихся клеток -"безработных", а также клеток -"пенсионеров", переставших заниматься общественно полезным трудом. Другая важная функция клеточной гибели - удаление клеток -"инвалидов" и клеток- "диссидентов" с серьезными нарушениями структуры или функции генетического аппарата. В частности, апоптоз - один из основных механизмов самопрофилактики онкологических заболеваний
Апоптоз играет главную роль как в развитии так и в гомеостазе. Клетки умирают от апоптоза в развивающемся эмбрионе в ходе морфогенеза или синантогенеза и во взрослых животных в ходе обновления тканей. Система программируемой клеточной смерти - существенный фактор иммунитета , поскольку гибель зараженной клетки может предотвратить распространение инфекции по организму. Формообразовательные процессы в онтогенезе , позитивная и негативная селекция Т- и В-лимфоцитов у животных, гипер-чувствительный ответ растений на вторжение патогена, осенний листопад - лишь несколько примеров программируемой клеточной смерти (апоптоза).
Многие инфекционные агенты выработали специальные меры для предотвращения преждевременной гибели зараженных клеток. Нарушения системы программируемой гибели клетки - причина серьезной патологии. Ослабление способности к апоптозу может вести к развитию злокачественных опухолей. Некоторые заболевания, в частности дегенеративные повреждения нервной системы, - результат избыточного апоптоза.
Исследуя нормальную и патологическую ткани, J.F.R. Кеrr с соавт. обнаружили, что умирающие клетки делятся на 2 категории. В сильно поврежденных тканях преобладают процессы некроза , которые затрагивают целые клеточные поля и характеризуются пассивной дегенерацией клеток с набуханием и фрагментацией органелл, разрушением мембран, лизисом клеток, выходом внутриклеточного содержимого в окружающую ткань и развитием воспалительного ответа. Некроз всегда обусловлен грубой патологией, его механизмы не требуют затрат энергии, и предотвратить его можно только, устранив причину повреждения.
Определенные клетки организма обладают уникальными сенсорами , называемыми рецепторами смерти , расположенными на поверхности клеток. Рецепторы смерти детектируют присутствие межклеточных сигналов смерти и в ответ на это быстро запускают внутриклеточный механизм апоптоза.
Поскольку физиологическая роль апоптоза очень существенна, нарушения этого процесса могут быть весьма вредными. Так, несвоевременный апоптоз определенных мозговых нейронов оказывает влияние на образование нарушений, таких как болезни Альтцгеймера и Паркинсона , в то время как неспособность делящихся клеток перейти к апоптозу после случившихся существенных нарушений ДНК способствует развитию рака.
В развитии апоптоза выделяют 3 стадии: сигнальную (индукторную), эффекторную и деградации (деструкции). Индукторами апоптоза могут быть как внешние (внеклеточные) факторы, так и внутриклеточные сигналы. Сигнал воспринимается рецептором и далее последовательно передается молекулам-посредникам (мессенджерам) различного порядка и достигает ядра, где происходит включение программы клеточного "самоубийства" путем активации летальных и/или репрессии антилетальных генов. В ядре регистрируются первые морфологические признаки апоптоза - конденсация хроматина с формированием его осмиофильных скоплений, прилежащих к ядерной мембране. Позже появляются инвагинации (вдавления) ядерной мембраны, и происходит фрагментация ядра. В основе деградации хроматина лежит ферментативное расщепление ДНК. Сначала образуются фрагменты, включающие 700, 200-250, 50-70 тыс. пар оснований, затем - фрагменты, содержащие 30-50 тыс. пар оснований. После реализации этого этапа процесс становится необратимым. Затем наступает межнуклеосомная дезинтеграция ДНК, т.е. разрывы нитей ДНК, находящихся между нуклеосомами. При этом образуются фрагменты, кратные по величине 180-190 пар оснований, что соответствует протяженности нити ДНК в пределах одной нуклеосомы. Отделившиеся фрагменты ядра, ограниченные мембраной , называют апоптотическими тельцами. В цитоплазме происходит расширение эндоплазматического ретикулума, конденсация и сморщивание гранул. Важнейшим признаком апоптоза является снижение трансмембранного потенциала митохондрий и выход в цитоплазму различных апоптогенных факторов (цитохрома с; прокаспаз 2, 3, 9; апоптоз-индуцирующего фактора). Именно нарушению барьерной функции митохондриальных мембран отводят ключевую роль в развитии многих типов апоптоза. Клеточная мембрана утрачивает ворсинчатость и образует пузыревидные вздутия. Клетки округляются и отделяются от субстрата. На поверхности клетки экспрессируются различные молекулы, распознаваемые фагоцитами - фосфосерин, тромбоспондин, десиалированные мембранные гликоконъюгаты, в результате чего происходит поглощение тела клетки другими клетками и его деградация в окружении лизосом фагоцитарных клеток Ярилин ea 2005
АПОПТОЗ: ВОЗМОЖНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ ЭТАПОВ
Сигналы апоптоза
Апоптоз является активным процессом саморазрушения клетки, требующим белкового синтеза. Это подтверждено in vivo в опытах по предупреждению смерти нейронов с помощью ингибиторов белкового синтеза Установлен благоприятный эффект инфузии ингибитора протеинового синтеза циклогексимида на выживаемость чувствительных нейронов CA1 зоны гиппокампа после преходящей ишемии мозга крыс. В то же время существуют данные о том, что апоптоз может проходить и в отсутствие синтеза протеинов de novo .
На первом этапе клетка получает "послание" о том, что она должна пожертвовать своей жизнью для благополучия организма. Это известие приходит из окружающей среды - либо от соседних клеток, либо от межклеточных веществ, твердых или жидких. Чтобы воспринять такое "послание", клетки имеют специальные органы чувств , которые называют рецепторами . Сигнальные молекулы и рецепторы подходят друг к другу, как ключ к замку. Информация передается внутрь клеток через различные рецепторы или сочетание рецепторов.
Информацией может являться и отсутствие специфического вещества в окружающей клетку среде. Хорошо известно, что в некоторых случаях молчащий телефон говорит очень о многом.
Существование апоптоза в безъядерных системах ( цитопластах - клетках, лишенных ядра) показывает, что наличие ядра не является обязательным для реализации процесса. Применительно к клеткам животных и человека апоптоз в большинстве случаев связан с протеолитической активацией каскада каспаз .
Однако апоптоз возможен и без участия каспаз : сверхсинтез белков- промоторов апоптоза Вах и Bak индуцирует апоптоз в присутствии ингибиторов каспаз .
Сигналы апоптоза передается внутрь клеток
В результате контакта сигнальных молекул с наружной частью белка-рецептора этот рецептор претерпевает структурные изменения. Структурная перестройка захватывает и внутриклеточную часть молекулы рецептора. Она может либо обладать определенной ферментативной активностью сама, либо быть тесно связана с некоторыми клеточными ферментами. Изменение структуры рецептора сказывается на работе этих ферментов, поэтому в результате контакта рецептора с внеклеточным веществом внутри клетки происходят разные биохимические изменения.
Часто речь идет об изменении концентрации ионов кальция , а также некоторых относительно мелких фосфорсодержащих органических соединений, относящихся к классу нуклеотидов (см циклические нуклеотиды ).
Активные соединения появляются и в результате гидролиза определенных липидов клеточной мембраны. В свою очередь, все это ведет к присоединению или отсоединению остатков фосфата от молекул белковых регуляторов (см фосфорилирование ), способных влиять на генетический аппарат клетки.
Фосфорилирование и дефосфорилирование, а также некоторые другие биохимические модификации меняют активность этих регуляторов.
» был впервые применен в 1972 г. Kerr, Wyllie et Currie для описания особой морфологической формы генетически запрограммированной гибели клеток, отличающейся от некроза.
Апоптоз является гомеостатическим механизмом, поддерживающим постоянство клеточной популяции в тканях, а также защитным механизмом при иммунных реакциях или при повреждении клеток при заболеваниях и при воздействии инфекционных агентов.
Апоптоз обусловлен процессами, которые вызывают активацию группы цистеиновых протеаз, называемых «каспазы», каскадный комплекс которых обуславливает в конечном итоге гибель клеток.
Поскольку клетки при апоптозе не освобождают свое содержимое в окружающие ткани и быстро фагоцитируются макрофагами, воспалительная реакция обычно отсутствует. Следует отметить, что пикноз и кариорексис не являются исключительным признаком апоптоза и могут быть частью цитоморфологического спектра при некрозе.
Ионизирующее излучение , химиопрепараты приводят к повреждению ДНК в клетках, что вызывает их гибель посредством в53-зависимого пути. Воздействие воспалительных агентов в малых дозах (гипоксия, радиация, повышение температуры) индуцирует апоптоз, вызывая некроз при воздействии в больших дозах.
При некрозе потеря клеточной мембраны приводит к освобождению цитоплазматического содержимого в окружающие ткани, посылая сигналы хемотаксиса, приводящего к клеточному воспалению. Хемотаксические факторы подразделяются на две категории: короткого и дальнего действия, формирующие навигационные сигналы в локальной области тканей, приводящие к миграции макрофагов из циркуляции.
Кроме того, радикально изменяется плазматическая мембрана апоптотических клеток: изменяется ее проницаемость, топология липидов с потерей фосфолипидной асимметрии, окисления и восстановления анионных фосфолипидов, фосфатидилсеринов с выходом их из клетки.
Изменяется также расположение углеводов на мембране, и различные белки (включая кальретикулин, аннексин 1), большие субъединицы инициирующего трансляцию фактора 3, ДНК, переносятся к поверхности апоптотических клеток и взаимодействуют (прямо или косвенно) с фагоцитами.
Такое изменение расположения макромолекул на поверхности апоптотических клеток является ключевым моментом во взаимодействии с фагоцитами. В процессе апоптоза происходит также потеря ингибиторных молекул (CD31 и CD47) с поверхности клеток (механизм «не ешь меня») с последующим взаимодействием апоптотических клеток с фагоцитами. Ниже схематически представлено большинство молекул, вовлеченных во взаимодействие между фагоцитами и апоптотичес-кими клетками (рис. 6).
Рис. 6. Молекулы, вовлеченные во взаимодействие между фагоцитами и апоптотическими клетками
Примечание: ABCA, АТФ-связывающиий кассетный транспортер A1; ACAMPs, апоптотические клеточно-связанные молекулярные партнеры; ASGP-R, рецептор асиалогликопротеина; 2GPI, 2 гликопротеин1; 2GPI-R, 2GPI-рецептор; интегрины, включая CR3 иCR4 ; BAI1, ангиогенный мозгово-специфический ингибитор; C1q, первый компонент комплемента; CHO, карбогидраты; CRP, С-реактивный белок; CRT, кальретикулин; CH3CR1, рецептор фракталькина; Del-1, эндотелиальный развивающий локус-1; FKN, фракталькин; GA, G-протеин-связанный LPC рецептор; Gas-6, фактор остановки роста-6; iC3b, инактивирующий комплемент фрагмент C3b; ICAM-3 (CD50), молекула-3 внутриклеточной адгезии; Lox-1, рецептор окисленного липопротеина низкой плотности; LPC, лизофосфатидил холин; MER, миелоидная эпителиальная репродуктивная тирозин киназа; MFG-E8, глобулин молочного жира эпидермального ростового фактора-8; Ox-PL, окисленный фосфолипид; P2Y2, G-протеин-связанный ядерный рецептор; PE, фосфатидилэтаноламин; PS, фосфатидилсерин; SAP, сывороточный амилоидный протеин; SHPS-1, гомолог2 доменсодержащего протеина субстрата-1 тирозин киназы; SR-AI, удаляющий рецептор (мусорщик) AI; SR-BI, удаляющий рецептор BI; TIM-1/4, молекулы Т-клеточного иммуноглобулина и муцин-содержащего домена; TSP-1, тромбоспондин-1.
Изменения клеток при апоптозе
Для клетки, подвергшейся апоптозу, характерно следующее.Сжатие клетки. Клетка уменьшается в размерах, цитоплазма уплотняется; органеллы, которые выглядят нормальными, располагаются более компактно. Предполагается, что нарушение формы и объема клетки происходит в результате активации в апоптотических клетках трансглютаминазы.
Этот фермент вызывает прогрессивное образование перекрестных связей в цитоплазматических белках, что приводит к формированию своеобразной оболочки под клеточной мембраной.
Конденсация хроматина
Это наиболее характерное проявление апоптоза. Хроматин конденсируется по периферии, под мембраной ядра, при этом образуются четко очерченные плотные массы разной формы и размеров.Ядро может разрываться на несколько фрагментов. Конденсация хроматина обусловлена расщеплением ядерной ДНК в местах, связывающих отдельные нуклеосомы, что приводит к образованию большого количества фрагментов, в которых число пар оснований составляет 180-200.
При электрофорезе фрагменты дают характерную картину лестницы (клеваж ДНК). Фрагментация ДНК в нуклеосомах происходит под действием кальций чувствительной эндонуклеазы.
Эндонуклеаза в некоторых клетках находится постоянно; в тимоцитах она активируется появлением в цитоплазме свободного кальция, а в других клетках синтезируется перед началом апоптоза.
Формирование апоптотических телец
В апоптотической клетке первоначально формируются глубокие впячивания поверхности с образованием полостей, что приводит к фрагментации клетки и формированию окруженных мембраной апоптотических телец, состоящих из цитоплазмы и плотно расположенных органелл с фрагментами ядра или без таковых.Апоптотические тельца быстро разрушаются в лизосомах макрофагов, а окружающие клетки либо мигрируют, либо делятся, чтобы заполнить освободившееся после гибели клетки пространство. Фагоцитоз апоптотических телец макрофагами или другими клетками активируется рецепторами на данных клетках: они захватывают и поглощают эти тельца.
В табл. 5 приводятся сравнительные данные по морфологическим изменениям при апоптозе и некрозе
Таблица 5. Морфологические изменения при апоптозе и некрозе

Геном человека содержит около 13 каспаз, их количество зависит от генетического полиморфизма. Каспазы присутствуют в цитоплазме в виде проэнзимов и активируются до полностью функциональных протеаз путем расщепления энзима на малую и большую субъединицы и дальнейшего отщепления от их N-концевых доменов.
Затем субъединицы собираются в тетрамер с двумя активными центрами. Расщепление прокаспаз могут осуществлять различные протеазы, в том числе и другие каспазы. По выполняемой каспазами функции их разделяют на две группы: инициаторные каспазы (8, 9 и 10) и эффекторные каспазы (3, 6, и 7). После того, как каспазы из первой группы активируют эффекторные каспазы, процесс апоптоза оказывается необратимым.
Расщепление каспазами ряда ключевых субстратов приводит к фрагментации ДНК и деструкции клетки. Для активации каспаз существует несколько путей, два из которых наиболее изучены и привлекают большое внимание в последнее время. Эти два пути апоптоза обычно обозначаются как внешний и внутренний путь.
Ниже представлено схематическое изображение разных путей апоптоза

Рис. 7. Схемы внешнего и внутреннего путей апоптоза, а также перфорин/гранзимного пути, который действует по каспазонезависимому пути. Результаты всех путей приводят к цитоморфологическим изменениям, включая сморщивание клеток, конденсацию хроматина, образование цитоплазматических и апоптотических телец, что в итоге приводит к фагоцитозу
апоптотических телец
Внутренний путь апоптоза (intrisic pathway)
При активации каспаз по внутреннему пути (intrinsic pathway) центром инициации апоптоза являются митохондрии. Стимуляция внутреннего пути продуцирует внутриклеточные сигналы, которые могут действовать как положительно, так и отрицательно.Негативные сигналы включают отсутствие факторов роста, гормонов и цитокинов, что ведет к нарушениям супрессии программы клеточной смерти, активируя апоптоз. Другие положительные стимулы включают радиацию, токсины, гипоксию, гипертермию, вирусные инфекции и свободные радикалы.
Все эти стимулы вызывают изменения внутренней мембраны митохондрий, результатом чего является открытие пор митохондриальной проницаемости, потеря митохондриального мембранного потенциала и освобождение двух наиболее больших групп проапоптотических протеинов. В митохондрии сходятся многие сигналы, вызывающие повреждение ДНК, нарушения микротрубочек, факторов роста, что вызывает освобождение из этих органелл в цитозол цитохрома с и других апоптогенных белков.
В цитозоле цитохром с связывается с белком, активирующим каспазы, апоптотическим протеазе -активирующим фактором 1 (Apafl). Apafl играет роль арматуры, на которой происходит аутокаталитический процессинг каспазы-9. В результате зависимого от гидролиза аденозинтрифосфата (АТФ) конформационного изменения Apaf1 приобретает способность связывать цитохром с.
Связав цитохром с, Apafl претерпевает дальнейшее конформационное изменение, способствующее его олигомеризации в гептаметрический комплекс и открывающее доступ каспазоизменяющего домена (CARD) Apafl для прокаспазы-9, которая также содержит CARD-домен.
В результате формируется мультипротеиновая структура, известная как «апоптосома». Связь Apafl с прокаспазой-9 обусловлена CARDs посредством гомотипического соединения (CARD-CARD). Активация апоптосомо-ассоциированной протеазы каспазы-9 инициирует протеолитический каскад, который активирует клеваж каспазы-9 и активирует прокаспазу-3.
Другая группа проапоптотических протеинов, апоптоз-индуцирующих факторов (AIF), эндонуклеазы G и CAD (каспазоактивируемая ДНКаза), освобождаются из митохондрий в процессе апоптоза. AIF фрагментирует ДНК и обуславливает конденсацию периферического ядерного хроматина.
Эндонуклеаза G перемещается в ядро, где расщепляет ядерный хроматин, образуя фрагменты олигонуклеосомальной ДНК. AIF и эндонуклеаза G действуют по каспазонезависимому пути. CAD последовательно освобождается из митохондрий и перемещается в ядро, где приводит к фрагментации олигонуклеосомальной ДНК и распространенной конденсации хроматина.
Контроль и регулирование данного митохондриального пути осуществляется белками семейства Bcl-2. Протеин гена-супрессора р53 играет критическую роль в регуляции белков семейства Bcl-2. Семейство белков Bcl-2 контролирует проницаемость митохондриальной мембраны и может действовать проапоптотически или антиапоптотически. Хотя геном человека содержит 25 членов этого семейства, только 6 из них являются антиапоптотическими.
Семейство белков Bcl-2 можно разделить на три основные группы:
1. Антиапоптогенные молекулы, такие как Bcl-2, Bcl-XL, Mcl-1, Bcl-W, Bfl-1, Bcl-B. Все они обладают антиапоптозной активностью, имеют четыре группы гомологичных последовательностей - ВН1, ВН2, ВН3 и ВН4 домены, хотя у некоторых из них домен ВН4 отсутствует. Эти молекулы представляют мембранные белки, находящиеся в митохондрии, эндоплазматическом ретикулуме и в ядерной мембране.
2. Проапоптогенные молекулы Bax, Bad, Bak, Mtd(Bok) и Diva имеют гомологичные последовательности ВН1, ВН2 и ВН3, а ВН4 домен у них отсутствует.
3. Проапоптогенные белки, содержащие только ВН3 домен: Bik, Bid, Bim, Hrk (DR5), Blk, Bnip3, Bnip3L.
Они в основном локализованы в цитозоле или связаны с цитоскелетом.
ВН1 -3 домены играют важную роль в формировании гетеро-и гомодимеров между проапоптогенными и антиапоптогенными членами семейства, и клеточные уровни этих димеров играют определяющую роль в судьбе клетки.
Гетеродимеризация происходит путем взаимодействия ВН3 домена проапоптогенного белка с гидрофобным комплексом, образованным ВН1, ВН2 и ВН3 доменами антиапоптогенных белков.
Домены ВН1, ВН2 и ВН4 необходимы для антиапоптогенной активности белка, в то время как ВН3 домен необходим для протоапоптогенной активности.
Функция белка Bcl-2 может быть дополнена возможностью посттрансляционной модификации с помощью фосфорилирования. Близкий ген, Bcl-x кодирует два белка, различающихся сплайсингом РНК, Bcl-xL и Bcl-xS. Так же как Bcl-2, белок Bcl-xL ингибирует апоптоз, в то время как белок Bcl-xS оказывает негативный эффект на функцию Bcl-2 и Bcl-xL.
Повышенная экспрессия генов этих белков может приводить к устойчивости к большинству вызывающих апоптоз стимулов, так как к этим белкам сходится множество путей апоптоза. Гиперэкспрессия некоторых антиапоптотических протеинов доказана при различных гематологических новообразованиях. Например, повышение уровня белка Bcl-2 в результате t(14; 18), вовлекающей ген BCL2, наблюдается в 80-90% случаев фолликулярной неходжкинской лимфомы.

Рис. 8. Схема апоптоза с участием всех ключевых факторов
Примерно 1/3 пациентов с диффузной В-крупноклеточной лимфомой имеют патологическое повышение уровня Bcl-2 (часто в ассоциации с t(14;18) или амплификацией гена), что коррелирует со снижением продолжительности жизни, несмотря на проведение комбинированной химиотерапии с включением ритуксимаба (анти-CD20 антител).
Большинство пациентов с ХЛЛ содержат повышенный уровень Bcl-2, ассоциированный с гипометилированием гена BCL2. В противоположность генетическим изменениям, активирующим антиапоптотические гены BCL2 и MCL1, при лейкозах и лимфомах с нестабильными микросателлитами часто происходят мутации, инактивирующие проапоптотический ген BAX.
Баланс между проапоптотическими и антиапоптотическими регуляторами апоптоза является основным механизмом, обеспечивая выживаемость длительно живущих клеток и замену ими короткоживущих клеток в различных тканях, включая костный мозг, тимус и периферические лимфоидные ткани. Дисбаланс этих протеинов в конечном итоге приводит к избирательным преимуществам в выживании клеток, что приводит к развитию новообразований.
Номенклатура основных белков внутреннего пути апоптоза приведена в табл. 6.
Таблица 6. Номенклатура основных белков внутреннего пути апоптоза

Внешний путь апоптоза (extrinsic pathway)
При внешнем пути апоптоза вначале происходит трансмембранное освобождение посредством фактора некроза опухоли (ФНО) рецептора смерти. Гибель клеток посредством внешнего пути апоптоза, в частности, химиорезистентных клеток, часто происходит при нарушениях по внутреннему пути, что дает преимущества при воздействии цитостатиков, ионизирующего излучения при выключении апоптоза клеток по митохондриальному пути.Семейство цитокинов ФНО состоит у человека из 18 членов. Некоторые из рецепторов семейства ФНО передают сигналы преимущественно для выживания клеток путем связывания внутриклеточного опухолевого рецептор-ассоциированного фактора (TRAF), семейства адаптерных белков. Блокирование этих рецепторов представляет новую стратегию в терапии лимфоидных опухолей. Другие члены семейства ФНО напрямую включают апопотоз, в частности, те, которые содержат «домен смерти» в их цитозольной части.
Стратегия для применения вызывающих апоптоз лиганд семейства ФНО включает: рекомбинантные лиганды, экспрессируемые только экстрацеллюлярной частью мембранных протеинов; моноклональные антитела, которые связывают рецепторы и включают апоптоз.
Подгруппа рецепторов семейства ФНО имеет цитоплазматический домен, состоящий из 80 аминокислот, именуемый «домен смерти» (DD), который при внутриклеточном взаимодействии с белками-адаптерами привязывает эти рецепторы к специфическим каспазам. Домен смерти играет основную роль в передаче сигнала смерти с поверхности клетки по внутриклеточному пути.
Связавшись с лигандом, рецепторы семейства ФНО образуют кластеры цитозольного DD на мембране, изменяя каспазосвязанный адаптерный протеин. Образующееся соединение адаптерного Fas-ассоциированного протеина с доменом смерти (FADD) состоит из DD и содержащего эффектор домена смерти (DED). DED в составе FADD связывает DED-содержащие прокаспазы (в частности, каспазы 8 и 10), формируя «смерть-индуцирующий сигнальный комплекс» (DISC), в результате чего происходит активация каспаз.
После активации каспазы-8 включается заключительная фаза апоптоза. Апоптоз, обусловленный рецептором смерти, может ингибироваться протеином c-FLIP (протеин, ингибирующий FLICE), который связывается с FADD и каспазой-8, делая их неэффективными.
Ниже представлена схема пути активации каспаз.

Рис. 9. Пути активации каспаз
Наличие дополнительных путей апоптоза включает:
1) путь апоптоза, индуцированный цитотоксическими лимфоцитами (CTL) и натуральными киллерами (NK), при котором сериновая протеаза гранзим В проникает внутрь клетки;
2) путь стресса эндоплазматического ретикулума (ER) с вовлечением каспазы-12;
3) р53-индуцированный путь, опосредoванный р53-индуцированным доменом смерти (PIDD), который связывает адапторный протеин ICH-1/протеин-3 (CED-3) домена смерти с доменом смерти, как активатором каспазы-2.
Номенклатура основных белков внешнего пути апоптоза приведена в табл. 7.
Таблица 7. Номенклатура основных белков внешнего пути апоптоза

Перфорин-гранзимный путь апоптоза (perforin-granzime pathway)
Цитотоксические Т-лимфоциты способны уничтожать клетки-мишени посредством внутреннего пути и FasL/FasR взаимодействия, что является основным способом апоптоза, вызываемого цитотоксическими лимфоцитами. Но они способны также осуществлять свой цитотоксический эффект в отношении опухолевых или инфицированных вирусом клеток посредством нового пути. Он осуществляется посредством секреции молекул перфорина.Полимеризуясь, перфорин образует в цитоплазматической мембране клетки-мишени трансмембранные каналы с последующим проникновением в клетки ФНО-в и гранзимов А и В - смеси сериновых протеаз. Гранзим В активирует прокаспазу-10 и может расщеплять ингибитор каспазактивируемой ДНКазы, а также использовать митохондриальный путь для амплификации сигнала смерти и вызывать освобождение цитохрома с.
Кроме того, гранзим В может напрямую активизировать каспазу-3, что может индуцировать заключительную фазу апоптоза. Таким образом, митохондриаль-ный путь и прямая активация каспазы-3 являются основными путями индуцированного гранзимом В уничтожения клеток.
Гранзим А также вызывает апоптоз, активируя каспазонезависимый путь апоптоза. Он расщепляет посредством активации ДНКазы продукт гена тумор-супрессора, вызывая апоптоз опухолевых клеток. Инактивация этого белка ведет к апоптозу вследствие блокирования восстановления ДНК и структуры хроматина.
Внешний и внутренний пути апоптоза заканчиваются в экзекутивной (исполнительной) фазе. Эта фаза начинается с активации экзекутивных («казнящих») каспаз, которые активируют цитоплазматические эндонуклеазы с деградацией ядерного материала, и активируют протеазы с последующей деградацией ядерных протеинов и протеинов цитоскелета. Каспаза-3 является наиболее важной экзекутивной каспазой и может активироваться любой каспазой (каспаза-8, каспаза-9 или каспаза-10).
Номенклатура основных белков экзекутивного пути апоптоза представлена в табл. 8.
Таблица 8. Номенклатура основных белков экзекутивного пути апоптоза

Е.В. Зуховицкая, А.Т. Фиясь
Введение
апоптоз смерть старение патологический
Апоптоз - физиологическая смерть клетки, представляющая собой своеобразную генетически запрограммированную самоликвидацию. Термин «апоптоз» в переводе с греческого означает «опадающий». Авторы термина дали такое название процессу запрограммированной смерти клеток потому, что именно с ним связано осеннее опадание увядших листьев. Кроме того, само название характеризует процесс как физиологический, постепенный и абсолютно безболезненный. У животных в качестве наиболее яркого примера апоптоза, как правило, приводят исчезновение хвоста у лягушки во время метаморфозы из головастика во взрослую особь. По мере взросления лягушонка хвост полностью исчезает, поскольку его клетки подвергаются постепенному апоптозу - запрограммированной смерти, и поглощению деструктированных элементов другими клетками. Явление генетически запрограммированной гибели клеток встречается у всех эукариотов (организмов, клетки которых имеют ядро). Прокариоты же (бактерии) имеют своеобразный аналог апоптоза. Можно сказать, что данный феномен характерен для всего живого, за исключением таких особых доклеточных форм жизни, как вирусы. Апоптозу могут подвергаться как отдельные клетки (как правило, дефектные), так и целые конгломераты. Последнее особенно характерно для эмбриогенеза. К примеру, опыты исследователей доказали, что благодаря апоптозу во время эмбриогенеза исчезают перепонки между пальцами на лапках у цыплят. Ученые утверждают, что у человека такие врожденные аномалии, как сросшиеся пальцы на руках и ногах, также возникают вследствие нарушения нормального апоптоза на ранних стадиях эмбриогенеза.
1. История открытия
Изучение механизмов и значения генетически программируемой клеточной смерти началось еще в шестидесятых годах прошлого века. Ученых заинтересовал тот факт, что клеточный состав большинства органов на протяжении жизни организма практически одинаков, а вот жизненный цикл различных типов клеток значительно отличается. При этом происходит постоянная замена многих клеток. Таким образом, относительное постоянство клеточного состава всех организмов поддерживается динамическим равновесием двух противоположных процессов - клеточной пролиферации (деление и рост) и физиологического отмирания отживших клеток. Авторство термина принадлежит британским ученым - Дж. Керру, Э. Уайли и А. Керри, которые впервые выдвинули и обосновали концепцию о принципиальном различии физиологической смерти клеток (апоптоз), и их патологической гибели (некроз). В 2002 году ученые из кембриджской лаборатории, биологи С. Бреннер, Дж. Салстон и Р. Хорвиц, получили Нобелевскую Премию по физиологии и медицине за раскрытие основных механизмов генетической регуляции развития органов и исследования программируемой клеточной смерти. Сегодня теории апоптоза посвящены десятки тысяч научных работ, раскрывающие основные механизмы его развития на физиологическом, генетическом и биохимическом уровнях. Ведется активный поиск его регуляторов. Особенно большой интерес представляют исследования, дающие возможность практического применения регуляции апоптоза при лечении онкологических, аутоиммунных и нейродистрофических заболеваний. В организме среднестатистического взрослого человека в результате апоптоза погибает ежедневно порядка 50-70 миллиардов клеток. Для среднестатистического ребёнка в возрасте от 8 до 14 лет число клеток, погибших путём апоптоза, составляет порядка 20-30 миллиардов в день. Суммарная масса клеток, которые на протяжении 1 года жизни подвергаются разрушению, эквивалентна массе тела человека. При этом восполнение утраченных клеток обеспечивается за счёт пролиферации - увеличения клеточной популяции путём деления.
Апоптоз человеческого лейкоцита
2. Механизм
Механизм развития апоптоза на сегодняшний день до конца не изучен. Доказано, что процесс может индуцироваться малыми концентрациями большинства веществ, вызывающих некроз. Однако в большинстве случаев генетически запрограммированная гибель клеток происходит при поступлении сигналов от молекул - клеточных регуляторов. В биохимическом механизме апоптоза выделяют 4 основных компонента: 1) CysAsp-протеазы илм каспазы; 2) так называемые «рецепторы серти» на поверхности клетки; 3) митохондрии и выходящий из них цитохром с и 4) специальные про-и антиапоптозные белки. Такие вторичные мессенджеры, как Са2+, активные формы кислорода (АФК) и окись азота (NO), также играют важную роль в апоптозе.
Каспазы (семейство CysAsp-протеаз) выполняют центральную роль в запуске апоптоза. У млекопитающих семейство каспаз состоит из 14 белков, постоянно синтезируемых практически во всех клетках в виде проферментов, активирующихся при апоптозе. Они принимают участие в развитии воспалительных процессов, а также, наряду с эффекторными каспазами, в пролиферации Т-лимфоцитов, терминальной дифференцировке эпителиальных клеток хрусталика и кератиноцитов.
Так называемые «рецепторы смерти». У млекопитающих апоптоз часто начинается с активации так называемых «индуцирующих смерть сигнальных комплексов» на плазматической мембране. Эти комплексы образуются при взаимодействии определенных внеклеточных лигандов - например, Fas или TNF (tumor necrosis factor) с белками семейства рецепторов фактора некроза опухолей (TNFR) на клеточной мембране, называемых «рецепторами смерти». При связывании лигандов они активируют каспазу-8, образуя «индуцирующий смерть сигнальный комплекс», содержащий «рецептор смерти», адаптеры TRADD (TNFR1-associated protein with death domain) или FADD (Fasassoccated protein with death domain) и профермент каспазы-8.
Митохондриям принадлежит центральная роль в осуществлении апоптоза у млекопитающих. Сигналы от рецепторов смерти или от поврежденных участков клетки сходятся на них, вызывая повышение проницаемости обеих мембран, снижение мембранного потенциала (??m) и высвобождение белков апоптоза - апоптоз-индуцирующего фактора (AIF), SMAC (second mitochondria derived activator of caspases) и некоторых прокаспаз - из межмембранного пространства.
Наряду со специфически апоптозными белками, в цитоплазму выходит цитохром c. Там он связывается с Apaf-1 (apoptotic protease activating factor-1) и формирует так называемый апоптосомный комплекс, инициирующий активацию каспазного каскада. С помощью Smac и Omi/HtrA2 (Omi stress regulated endopeptidase/high temperature requirement protein A2) цитохром с запускает Apaf-1 зависимую активацию каспазы-9. Каспаза-9 активирует каспазы-3 и -7 (рисунок); те в свою очередь расщепляют различные белки, приводя к появлению биохимических и морфологических признаков аоптоза.
3. Фазы апоптоза
Стадии апоптоза
Различают три физиологические фазы апоптоза:
1. Сигнальная (активация специализированных рецепторов).
Инициация апоптоза может происходить посредством внешних (внеклеточных) или внутриклеточных факторов. Например, в результате гипоксии, гипероксии, субнекротического поражения химическими или физическими агентами, перекрёстного связывания соответствующих рецепторов, нарушения сигналов клеточного цикла, удаления факторов роста и метаболизма и т.д. Несмотря на разнообразие инициирующих факторов, выделяются два основных пути передачи сигнала апоптоза: рецептор- зависимый (внешний) сигнальный путь с участием рецепторов гибели клетки и митохондриальный(собственный) путь.
Рецептор-зависимый сигнальный путь
Процесс апоптоза часто (например, у млекопитающих) начинается с взаимодействия специфических внеклеточных лигандов с рецепторами клеточной гибели, экспрессированными на поверхности клеточной мембраны. Рецепторы, воспринимающие сигнал апоптоза, относятся к суперсемейству TNF-рецепторов (англ. tumor necrosis factor receptor или кратко TNFR - «рецептор фактора некроза опухолей»). Наиболее изученными рецепторами смерти, для которых описана и определена роль в апоптозе, являются CD95 (также известный как Fas или APO-1) и TNFR1 (также называемый p55 или CD120a). К дополнительным относятся CARI, DR3 (англ. death receptor 3 - «рецептор смерти 3»), DR4 и DR5.
Все рецепторы смерти представляют собой трансмембранные белки, характеризующиеся наличием общей последовательности из 80 аминокислот в цитоплазматическом домене. Данная последовательность называется доменомсмерти (англ. death domain или кратко DD) и является необходимой для трансдукции сигнала апоптоза.Внеклеточные участки рецепторов смерти взаимодействуют с тримерами лигандов (CD95L, TNF, Apo3L, Apo2L и т.п.). Тримеры лигандов в результате взаимодействия тримеризуют рецепторы смерти (то есть «сшивают» 3 молекулы рецептора). Активированный таким образом рецептор взаимодействует с соответствующим внутриклеточным адаптером (или адаптерами). Для рецептора CD95 (Fas/APO-1) адаптером является FADD (от англ. Fas-associated DD-protein - «белок, взаимодействующий с доменом смерти Fas-рецептора»). Для рецепторов TNFR1 и DR3 адаптером является TRADD (от англ. TNFR1-associated DD-protein - «белок, взаимодействующий с доменом смерти TNFR1-рецептора»).
Адаптер, ассоциированный с рецептором смерти, вступает во взаимодействие с эффекторами - пока ещё неактивными предшественниками протеаз из семейства инициирующих каспаз - с прокаспазами. В результате цепочки взаимодействия «лиганд-рецептор-адаптер-эффектор» формируются агрегаты, в которых происходит активация каспаз. Данные агрегаты именуются апоптосомами, апоптозными шаперонами или сигнальными комплексами, индуцирующими смерть (от англ. DISC - death-inducing signaling complex - «сигнальный комплекс, индуцирующий смерть»). Примером апоптосомы может служить комплекс FasL-Fas-FADD-прокаспаза-8, в котором активируется каспаза-8.
Рецепторы смерти, адаптеры и эффекторы взаимодействуют между собой сходными по структуре доменами: DD, DED, CARD. DD (от англ. death domain - «домен смерти») участвует во взаимодействии рецептора Fas с адаптером FADD и во взаимодействии рецепторов TNFR1 или DR3 с адаптером TRADD. Посредством домена DED (от англ. death-effector domain - «домен эффектора смерти») осуществляется взаимодействие адаптера FADD с прокаспазами?8 и?10. Домен CARD (от англ. caspase activation and recruitment domain - «домен активации и рекрутирования каспазы») участвует во взаимодействии адаптера RAIDD с прокаспазой-2.
Посредством рецепторов смерти могут быть активированы три инициирующие каспазы: ?2; ?8 и?10. Активированные инициирующие каспазы далее участвуют в активации эффекторных каспаз.
Митохондриальный сигнальный путь
Большинство форм апоптоза у позвоночных реализуется по митохондриальному пути, а не через рецепторы клеточной гибели. Митохондриальный сигнальный путь апоптоза реализуется в результате выхода апоптогенных белков из межмембранного пространства митохондрий в цитоплазму клетки. Высвобождение апоптогенных белков, предположительно, может осуществляться двумя путями: за счёт разрыва митохондриальной мембраны или же путём открытия высокопроницаемых каналов на внешней мембране митохондрий.
Ключевым событием митохондриального пути апоптоза является повышение проницаемости наружной мембраны митохондрий (англ. Mitochondrial Outer Membrane Permeabilization, MOMP). Существенную роль в повышении MOMP играют апоптотические Bcl-2 белки - Bax и Bak. Они встраиваются в наружную мембрану митохондрий и олигомеризуются. При этом, вероятно, нарушается целостность внешней мембраны митохондрий, по неизвестному пока механизму. При повышении MOMP из межмембранного пространства митохондрий в цитозоль высвобождаются растворимые белки, участвующие в апоптозе: цитохром c - белок с молекулярной массой 15 кДа; прокаспазы?2, ?3 и?9; AIF (от англ. apoptosis inducing factor - «фактор индуцирующий апоптоз») - флавопротеин с молекулярной массой 57 кДа.
Разрыв внешней мембраны митохондрий объясняется увеличением объема митохондриального матрикса. Данный процесс связывают с раскрытием пор митохондриальной мембраны, приводящим к снижению мембранного потенциала и высокоамплитудному набуханию митохондрий вследствие осмотического дисбаланса. Поры диаметром 2,6-2,9 нм способны пропускать низкомолекулярные вещества массой до 1,5 кДа. Раскрытие пор стимулируют следующие факторы: неорганический фосфат; каспазы; SH-реагенты; истощение клеток восстановленным глутатионом; образование активных форм кислорода; разобщение окислительного фосфорилирования протонофорными соединениями; увеличение содержания Ca2+ в цитоплазме; воздействие церамида; истощение митохондриального пула АТФ и др.
Цитохром c в цитоплазме клетки участвует в формировании апоптосомывместе с белком APAF-1 (от англ. Apoptosis Protease Activating Factor-1 - «активирующий фактор апоптотической протеазы-1»). Предварительно, APAF-1 претерпевает конформационные изменения в результате реакции, протекающей с затратой энергии АТФ. Предполагается, что трансформированный APAF-1 приобретает способность связывать цитохром c . К тому же открывается доступ CARD-домена APAF-1 для прокаспазы-9. В итоге происходит олигомеризация 7 субъединиц трансформированного белка APAF-1 с участием цитохрома c и прокаспазы-9. Так образуется апоптосома, активирующая каспазу-9. Зрелая каспаза-9 связывает и активирует прокаспазу-3 с образованием эффекторной каспазы-3. Высвобождающийся из межмембранного пространства митохондрий флавопротеин AIF является эффектором апоптоза, действующим независимо от каспаз.
2. Эффекторная (т.е. формирование из разнородных эффекторных сигналов единого пути апоптоза, и запуск каскада сложных биохимических реакций).
В течение эффекторной фазы различные инициирующие пути конвертируются в один (или несколько) общий путь апоптоза. Как правило, происходит активация каскада белков-эффекторов и регулирующих их белков-модуляторов. Основными эффекторами апоптоза являются каспазы. В процессе активации они запускают каспазный каскад: сложно переплетённые цепочки взаимодействий инициирующих и эффекторных каспаз:
Помимо каспазсуществуют и другие эффекторы апоптоза. Например, флавопротеин AIF, высвобождающийся из межмембранного пространства митохондрий, действует по независимому от каспаз пути. Попадая в клеточное ядро, AIF вызывает конденсацию хроматина и активирует эндонуклеазы, которые участвуют в фрагментации ДНК. На основании экспериментальных данных установлено, что апоптоз, протекающий в присутствии AIF, не предотвращается ингибитором каспаз. В качестве эффекторов апоптоза также рассматриваются кальпаины - представители семейства цитозольных Ca2+-активируемых цистеиновых протеаз. Их роль в апоптозе пока слабо охарактеризована.
Деградационная (фаза экзекуции или деструкции).
Условно деградацию погибающей клетки можно разделить на три последовательных фазы: высвобождения, блеббинга и конденсации. Деградация большинства клеток начинается с высвобождения прикреплений внеклеточного матрикса и реорганизации фокальной адгезии. Внутри погибающей клетки деполимеризуются микротрубочки цитоскелета. Внутриклеточные актиновые микрофиламенты реорганизуются в связанные с мембраной периферийные (кортикальные) кольцевые пучки. В итоге клетка приобретает округлую форму. Следующая за высвобождением, стадия блеббинга, характеризуется сокращением периферийных актиновых колец. В результате сокращений клеточная мембрана образует вздутия, клетка как бы «кипит». Процесс блеббинга энергозависим и требует большого количества АТФ. Фаза блеббинга в нормальных условиях завершается примерно через час. В итоге клетка фрагментируется на маленькие апоптотические тела, либо целиком конденсируется, округляясь и уменьшаясь в размерах.
Роль белка р53
В нормальных клетках белок p53, как правило, находится в неактивной, латентной форме. Активация p53 происходит в ответ на повреждения ДНК, вызванные ультрафиолетовым или гамма-излучением, гиперэкспрессией онкогенов, вирусной инфекцией, оксидативным стрессом, гипо- и гипертермией и др.Активированный p53 координирует процесс репарации ДНК, а также регулирует транскрипцию ряда генов-активаторов апоптоза в случае необратимых повреждений ДНК или нарушений регуляции клеточного цикла. К тому же имеются указания на то, что p53 принимает участие в запуске апоптоза путём стимуляции рецепторов смерти, путём взаимодействия с промотором апоптоза - Bax, путём активации p53-зависимого модулятора апоптоза PUMA (англ. p53 upregulated modulator of apoptosis), который блокирует действие Bcl-2.Повышение уровня p53 в ответ на повреждения ДНК вызывает апоптоз, например, в клетках кожи, в тимоцитах, в клетках кишечного эпителия.
4. Роль апоптоза в процессах старения
Предположение о роли апоптотической гибели в процессах старения было высказано ещё в 1982 году. Со временем выяснилось, что различные виды возрастзависимой дисрегуляции апоптоза присущи многим типам клеток. Например, в стареющем организме повышается чувствительность к индукции апоптоза для следующих типов клеток: гепатоцитов, кардиомиоцитов, макрофагов, мегакариоцитов, нейронов, ооцитов, спленоцитов, T-лимфоцитов, хондроцитов, эндотелиоцитов. Но в то же время, для фибробластовнаблюдается обратная тенденция к снижению чувствительности к апоптозу, а для кератиноцитов данная чувствительность не изменяется.
К настоящему времени имеются, как минимум, две точки зрения на связь апоптоза с процессами старения. Согласно одной из версий нормальные (гомеостатические) апоптотические процессы могут участвовать в развитии возрастных патологий и фенотипов старения. К примеру, с апоптотической гибелью постмитотических клеток (кардиомиоцитов, нейронов) связаны процессы старения сердечной мышцы или развитие возрастных нейродегенеративных патологий. Старение иммунной системы также связывают с программируемой гибелью различных типов лейкоцитов в результате возрастных изменений в соотношении про- и антиапоптозных факторов. Возрастная хрящевая дегенерация кореллирует с повышением уровня апоптоза хондроцитов в суставных хрящах у мышей и крыс, а также в межпозвоночных дисках при старении у человека. Согласно другой точке зрения накопление стареющих клеток в тканях объясняется возрастной резистентностью к апоптозу. В качестве примера, рассматривается устойчивость стареющих фибробластов к апоптозу, приводящая в итоге к преждевременному старению нормальных фибробластов и, возможно, к нарушению функций соединительной ткани.
5. Патология, связанная с усилением апоптоза
Одной из групп заболеваний, связанных с усилением апоптоза, являются патологии системы крови. Чаще всего патологические процессы развиваются в результате гибели посредством апоптоза костномозговых клеток-предшественников. Причиной их гибели является недостаточность факторов выживания. Данный тип патологии приводит к развитию апластической анемии; анемии при дефиците железа, фолатов, витамина B12; талассемии; тромбоцитопении; лимфопении; нейтропении; панцитопении. Повышенная готовность к развитию апоптоза Т-лимфоцитов обнаружена при мультицентрической болезни Кастелмана.
Прогрессия некоторых инфекционных заболеваний может быть связана не только с подавлением, но и наоборот, с усилением апоптоза. Индукторами программируемой клеточной гибели при этом служат бактериальные эндо- и экзотоксины. Массовый апоптоз развивается при сепсисе. Гибель лимфоцитов путём апоптоза находится в положительной корреляции с быстрой прогрессией СПИДа.
Отдельную группу патологии составляют заболевания нервной системы, обусловленные атрофией определённых участков нервной ткани в результате апоптоза. Примерами таких заболеваний могут служить боковой амиотрофический склероз, болезнь Альцгеймера, спинальная мышечная атрофия и др.
Апоптоз является преобладающей формой гибели миоцитов в ранний период развития инфаркта. На основе экспериментальных данных было выявлено, что программируемая гибель кардиомиоцитов может быть обусловлена гипоксией, ишемией, перегрузкой клетки кальцием, воспалением, токсинами. В процессе токсического (в том числе и алкогольного) гепатита основная роль также отводится апоптозу.
Ряд патологических процессов, обусловленных усилением апоптоза, индуцируется внешними апоптогенными факторами. Апоптоз прогрессирует под воздействием ионизирующей радиации.При этом преимущественно гибнут лимфоидные клетки и развивается иммунная недостаточность. Аналогичный эффект дают многие химиотерапевтические препараты, используемые при лечении опухолей, а также гормоны, применяемые при лечении различных заболеваний.
6. Другие формы ПКС (программируемой клеточной смерти)
·Аутофагию
·Некроз
Термином « Аутофагия» (Autophagy, от греческих слов: «Авто», означающими само- и «phagein означающее «поглощать») обозначают поглощение и переваривание в лизосомах «состарившихся» или поврежденных молекул или органелл собственной клетки. Аутофагия - необходимая часть обновления молекул и органелл клетки (вместе с образованием новых молекул и органелл). Внутриклеточный материал сначала включается в пузырьки, образуемые мембранами эндоплазматического ретикулума, а затем эти пузырьки сливаются с лизосомами.В каждой клетке печени за сутки разрушается около 100 митохондрий С/20 часть всех митохондрий).
При воспалительных процессах мембранные структуры клеток повреждаются, в том числе и мембраны лизосом. Лизосомные ферменты освобождаются и переваривают клетку; этот процесс может способствовать образованию язв. Разрушение соединительнотканного матрикса при таких заболеваниях, как ревматоидный артрит, миодистрофия, инфаркт миокарда, связано с освобождением лизосомальных ферментов. С другой стороны, гетерофагия и аутофагия участвуют в заживлении ран и воспалительных повреждений тканей, удаляя погибшие клетки или фрагменты клеток. Одна из важных функций эндоцитоза и лизосом связана с регуляцией количества рецепторов, экспонированных на поверхности клетки.
Некроз (от греч. ?????? - мёртвый), или омертве?ние - это патологический процесс, выражающийся в местной гибели ткани в живом организме в результате какого-либо экзо- или эндогенного её повреждения.
Некроз проявляется в набухании, денатурации и коагуляции цитоплазматических белков, разрушении клеточных органелл и, наконец, всей клетки. Наиболее частыми причинами некротического повреждения ткани являются: прекращение кровоснабжения (что может приводить к инфаркту, гангрене) и воздействие патогенными продуктами бактерий или вирусов (токсины, белки, вызывающие реакции гиперчувствительности, и др.)
7. Отличия между некрозом и апоптозом
Отличия апоптоза от некроза связаны с различиями в их встречаемости, биохимическими, генетическими, морфологическими и клиническими реакциями. Главным отличием апоптоза от некроза является то, что апоптоз распространяется исключительно на отдельные клетки или их совокупности, в то время как некроз может уничтожить территорию начиная от части клетки вплоть до органа.
Апоптоз происходит в клетках при определённых генетических событиях, которые во многом ещё недостаточно проанализированы. При апоптозе увеличивается экспрессия генов, ответственных запролиферацию и дифференцировку клеток из совокупности клеточных онкогенов (c-fos, c-myc, c-bcl-2) и антионкогенов (p53). Активация клеточных онкогенов должна привести к увеличению пролиферации клеток, однако при параллельной активации антионкогена р53 происходит апоптоз. Описанные взаимоотношения между генами показывают возможность регуляции процессов пролиферации и гибели клеток, построенной в генетическом аппарате клеток. В связи с тем, что взаимодействия между генами происходят с помощью их белковых соединений, в момент апоптоза в клетке увеличивается белковый синтез. Ингибирование этого процесса может предотвращать апоптоз.
Морфологические отличия апоптоза от некроза.Эти отличия касаются в основном ультраструктурных перестроек. Но это не значит, что апоптоз невозможно наблюдать на светооптическом уровне. При световой микроскопии клетки в состоянии апоптоза и их фрагменты (апоптозные тельца) отличаются небольшими размерами, сравнимыми с размерами лимфоцитов, с высоким ядерно-цитоплазматическим соотношением, округлыми контурами и конденсированными хроматином и цитоплазмой. Существенным отличием является также отсутствие воспалительной реакции на апоптоз.
Ультраструктурные отличия апоптоза от некроза.Существуют следующие ультраструктурные отличия. - Потеря специализированных структур клеточной поверхности - микроворсинок, межклеточных контактов. Клетка приобретает округлую форму и теряет связь с соседними клетками. В отличие от некроза речь идет всегда об изменениях в отдельных клетках.
- Размеры клеток уменьшаются в связи с конденсацией цитоплазматических органелл; изменяется также и форма клетки. Часто клетка расщепляется на несколько апоптозных телец, каждое из которых имеет свой фрагмент ядра, ограниченный двухконтурной ядерной мембраной, и индивидуальный набор орга-нелл.
В отличие от некроза при апоптозе имеется сохранность и интегративность органелл. Митохондрии не набухают, в них не происходит разрыва внутренней мембраны. Характерными для апоптоза являются такие ультраструктурные изменения, как агрегация рибосом в полукристаллоидные структуры, появление пучков микрофиламентов под цитолеммой, расположенных параллельно мембране. Почти всегда наблюдается кратковременная дилатация агранулярной эндоплазматической сети с формированием пузырей, наполненных жидкостью, которые выводятся из клетки. При изучении в сканирующем электронном микроскопе поверхность клетки приобретает кратерообразные выпячивания. - Наиболее яркое отличие апоптоза от некроза связано с изменениями ядерного хроматина, который конденсируется под кари-олеммой в виде полусфер и глыбок. В ядре обнаруживаются ос-миофильные тельца, сформированные транскрипционными комплексами, поступающими из ядрышек. Ядро меняет свою форму, становится изрезанным, фрагментирустся, ядерные поры концентрируются только в участках, где отсутствует маргинации хроматина.
Клетка в состоянии апоптоза становится объектом фагоцито-за для соседних паренхиматозных и стромальных клеток в прежде всего для макрофагов. Фагоцитоз происходит настолько быстро, что в условиях in vivo апоптозные клетки сохраняются лини, и течение нескольких минут, что затрудняет их наблюдение.
Заключение
В генетическом аппарате каждой клетки многоклеточного организма имеется специальная программа, которая при определенных обстоятельствах может привести клетку к гибели. При нормальном развитии эта программа направлена на удаление избыточно образовавшихся клеток - «безработных», а также клеток - «пенсионеров», переставших заниматься общественно полезным трудом. Другая важная функция клеточной гибели - удаление клеток - «инвалидов» и клеток «диссидентов» с серьезными нарушениями структуры или функции генетического аппарата. В частности, апоптоз - один из основных механизмов самопрофилактики онкологических заболеваний
Система программируемой клеточной смерти - существенный фактор иммунитета, поскольку гибель зараженной клетки может предотвратить распространение инфекции по организму. Другое дело, что некоторые инфекционные агенты выработали специальные меры для предотвращения преждевременной гибели зараженных клеток. Нарушения системы программируемой гибели клетки - причина серьезной патологии. Ослабление способности к апоптозу может вести к развитию злокачественных опухолей. Некоторые заболевания, в частности дегенеративные повреждения нервной системы, - результат избыточного апоптоза.
Воздействие на программу клеточной гибели - перспективное направление лекарственного лечения. Так, одна из важных задач противораковой терапии - стимуляция апоптозной системы. В других случаях задача врача, наоборот, предотвратить вредное для организма клеточное самоубийство. Таким образом, какие-то компоненты каждой клетки по праву могли бы нести микроскопическое изображение черепа со скрещенными костями. Однако следует признать, что наличие такого смертельного механизма - обстоятельство не только необходимое, но в конечном итоге крайне благоприятное. Без системы программируемой клеточной гибели мы с вами не могли бы появиться на свет такими, какими мы рождаемся. И поддержание порядка в наших организмах в течение дальнейшей жизни в значительной степени обеспечивается именно способностью наших клеток к программируемой смерти.
Список использованной литературы
1.А.В. Гордеева, Ю.А. Лабас, Р.А Звягильская: «АПОПТОЗ ОДНОКЛЕТОЧНЫХ ОРГАНИЗМОВ: МЕХАНИЗМЫ И ЭВОЛЮЦИЯ» Обзор. Институт биохимии им. А.Н. Баха РАН, Москва, 2004 год.
2.Анисимов В.Н.: «Молекулярные и физиологические механизмы старения» в 2 Т., СПб, 2008, 2-ое издание дополненное и переработанное.
.В.И. Агол: «Генетически запрограммированная смерть клетки», Московский Государственный университет им. М.В. Ломоносова, Соровский образовательный журнал №6, 2006 год.
.Бра М. Митохондрии в программированной гибели клетки: различные механизмы гибели / М. Бра, Б. Квинан, С.А. Сузин // Биохимия. 2005. - Т.70. - №2.
.Лушников Е.Ф. Гибель клетки (апоптоз) / Е.Ф. Лушников, А.Ю. Абросимов. М.: Медицина, 2001. - 192 с.
.#"justify">.#"justify">.#"justify">9.#"justify">10.#"justify">.http://tsitologiya.cytspb.rssi.ru/49_5/shirokova.pdf
Репетиторство
Нужна помощь по изучению какой-либы темы?
Наши специалисты проконсультируют или окажут репетиторские услуги по интересующей вас тематике.
Отправь заявку
с указанием темы прямо сейчас, чтобы узнать о возможности получения консультации.
Процесс, при котором клетка может убивать сама себя, называется запрограммированной клеточной гибелью (ЗГК). Этот механизм имеет несколько разновидностей и играет важнейшую роль в физиологии различных организмов, особенно многоклеточных. Самой часто встречающейся и хорошо изученной формой ЗГК является апоптоз.
Что такое апоптоз
Апоптоз - это контролируемый физиологический процесс самоуничтожения клетки, характеризующийся поэтапным разрушением и фрагментацией ее содержимого с формированием мембранных пузырьков (апоптозных телец), впоследствии поглощаемых фагоцитами. Этот генетически заложенный механизм активируется под воздействием определенных внутренних или внешних факторов.
При таком варианте гибели клеточное содержимое не выходит за пределы мембраны и не вызывает воспаление. Нарушения в регуляции апоптоза приводят к серьезным патологиям, таким как неконтролируемые клеточные деления или дегенерация тканей.
Апоптоз представляет собой лишь одну из нескольких форм запрограммированной гибели клетки (ЗГК), поэтому отождествлять эти понятия ошибочно. К известным видам клеточного самоуничтожения относят также митотическую катастрофу, аутофагию и программированный некроз. Другие механизмы ЗГК пока не изучены.
Причины апоптоза клеток
Причиной запуска механизма запрограммированной клеточной гибели могут быть как естественные физиологические процессы, так и патологические изменения, вызванные внутренними дефектами или воздействием внешних неблагоприятных факторов.
В норме апоптоз уравновешивает процесс деления клеток, регулируя их количество и способствуя обновлению тканей. В таком случае причиной ЗГК служат определенные сигналы, входящие в систему контроля гомеостаза. С помощью апоптоза уничтожаются одноразовые или выполнившие свою функцию клетки. Так, повышенное содержание лейкоцитов, нейтрофилов и других элементов клеточного иммунитета по окончании борьбы с инфекцией устраняется именно за счет апоптоза.
Запрограммированная гибель является частью физиологического цикла репродуктивных систем. Апоптоз задействован в процессе оогенеза, а также способствует гибели яйцеклетки при отсутствии оплодотворения.
Классическим примером участия апоптоза клеток в жизненном цикле вегетативных систем является осенний листопад. Сам термин происходит от греческого слова apoptosis, что буквально переводится как "опадание".
Апоптоз играет важнейшую роль в эмбриогенезе и онтогенезе, когда в организме сменяются ткани и атрофируются определенные органы. Примером могут служить исчезновение перепонок между пальцами конечностей некоторых млекопитающих или отмирание хвоста при метаморфозе лягушки.
Апоптоз может быть спровоцирован накоплением дефектных изменений в клетке, возникших в результате мутаций, старения или ошибок митоза. Причиной запуска ЗГК могут быть неблагоприятная среда (недостаток питательных компонентов, дефицит кислорода) и патологические внешние воздействия, опосредованные вирусами, бактериями, токсинами и т. д. При этом если повреждающий эффект слишком интенсивен, то клетка не успевает осуществить механизм апоптоза и погибает в результате развития патологического процесса - некроза.

Морфологические и структурно-биохимические изменения клетки во время апоптоза
Процесс апоптоза характеризуется определенным набором морфологических изменений, которые с помощью микроскопии можно наблюдать в препарате ткани in vitro.

К основным признакам, характерным для апоптоза клеток, относят:
- перестраивание цитоскелета;
- уплотнение клеточного содержимого;
- конденсацию хроматина;
- фрагментацию ядра;
- уменьшение объема клетки;
- сморщивание контура мембраны;
- образование пузырьков на клеточной поверхности,
- деструкцию органоидов.
У животных эти процессы завершаются образованием апоптоцитов, которые могут быть поглощены как макрофагами, так и соседними клетками ткани. У растений формирования апоптозных телец не происходит, а после деградации протопласта сохраняется остов в виде клеточной стенки.

Помимо морфологических изменений, апоптоз сопровождается рядом перестроек на молекулярном уровне. Происходит повышение липазной и нуклеазной активностей, которые влекут за собой фрагментацию хроматина и многих белков. Резко увеличивается содержание сАМФ, изменяется структура клеточной мембраны. В растительных клетках наблюдается образование гигантских вакуолей.
Чем апоптоз отличается от некроза

Главное различие между апоптозом и некрозом заключается в причине клеточной деградации. В первом случае источником разрушения служат молекулярные инструменты самой клетки, которые работают под строгим контролем и требуют затрат энергии АТФ. При некрозе происходит пассивное прекращение жизнедеятельности из-за внешнего повреждающего воздействия.
Апоптоз - это естественный физиологический процесс, сконструированный таким образом, чтобы не вредить окружающим клеткам. Некроз - это неконтролируемое патологическое явление, возникающее в результате критических повреждений. Поэтому неудивительно, что механизм, морфология и последствия апоптоза и некроза во многом противоположны. Однако имеются и общие черты.
В случае повреждения клетки запускают механизм запрограммированной гибели в том числе для того, чтобы не допустить некротического развития. Однако недавние исследования показали, что существует иная непатологическая форма некроза, которую также отнесли к ЗГК.
Биологическое значение апоптоза
Несмотря на то что апоптоз приводит к клеточной гибели, его роль для поддержания нормальной жизнедеятельности всего организма очень велика. Благодаря механизму ЗГК осуществляются следующие физиологические функции:
- поддержание баланса между пролиферацией и смертью клеток;
- обновление тканей и органов;
- устранение дефектных и "старых" клеток;
- защита от развития патогенного некроза;
- смена тканей и органов при эмбрио- и онтогенезе;
- удаление ненужных элементов, выполнивших свою функцию;
- устранение клеток, нежелательных или опасных для организма (мутантных, опухолевых, зараженных вирусом);
- предотвращение развития инфекции.
Таким образом, апоптоз является одним из способов поддержания клеточно-тканевого гомеостаза.

Этапы клеточной смерти
То, что происходит с клеткой при апоптозе, является результатом сложной цепочки молекулярных взаимодействий между различными ферментами. Реакции проходят по типу каскада, когда одни белки активируют другие, способствуя постепенному развитию сценария гибели. Этот процесс можно разделить на несколько этапов:
- Индукция.
- Активация проапоптических белков.
- Активация каспаз.
- Разрушение и перестройка клеточных органелл.
- Формирование апоптоцитов.
- Подготовка клеточных фрагментов к фагоцитозу.
Синтез всех компонентов, необходимых для запуска, реализации и контроля каждого этапа заложен генетически, почему апоптоз и называют запрограммированной гибелью клетки. Активация этого процесса находится под строгим контролем регуляторных систем, включающих в том числе и различные ингибиторы ЗГК.
Молекулярные механизмы апоптоза клетки
Развитие апоптоза обуславливается совокупным действием двух молекулярных систем: индукционной и эффекторной. Первый блок отвечает за контролируемый запуск ЗГК. В него входят так называемые рецепторы смерти, Cys-Asp-протеазы (каспазы), ряд митохондриальных компонентов и проапоптических белков. Все элементы индукционной фазы можно разделить на тригеры (участвуют в индукции) и модуляторы, обеспечивающие трансдукцию сигнала смерти.
Эффекторную систему составляют молекулярные инструменты, обеспечивающие деградацию и перестройку клеточных компонентов. Переход между первой и второй фазами осуществляется на этапе протеолитического каспазного каскада. Именно за счет компонентов эффекторного блока происходит гибель клетки при апоптозе.
Факторы апоптоза
Структурно-морфологические и биохимические изменения при апоптозе осуществляются определенным набором специализированных клеточных инструментов, среди которых наиболее важными являются каспасы, нуклеазы и мембранные модификаторы.
Каспазы - группа ферментов, разрезающих пептидные связи по остаткам аспарагина, фрагментируя белки на крупные пептиды. До начала апоптоза присутствуют в клетке в неактивном состоянии из-за ингибиторов. Главной мишенью каспаз являются ядерные белки.
Нуклеазы - ответственны за разрезание молекул ДНК. Особо важна в развитии апоптоза активная эндонуклеаза CAD, разрывающая участки хроматина в областях линкерных последовательностей. В результате образуются фрагменты длиной 120-180 нуклеотидных пар. Комплексное воздействие протеолитических каспаз и нуклеаз приводит к деформации и фрагментации ядра.

Модификаторы клеточной мембраны - нарушают асимметричность билипидного слоя, превращая его в мишень для фагоцитирующих клеток.
Ключевая роль в развитии апоптоза принадлежит каспазам, которые поэтапно активируют все последующие механизмы деградации и морфологической перестройки.
Роль каспаз в клеточной гибели
Семейство каспаз включает 14 белков. Часть из них не задействована в апоптозе, а остальные подразделяются на 2 группы: инициаторные (2, 8, 9, 10, 12) и эффекторные (3, 6 и 7), которые иначе называются каспазами второго эшелона. Все эти белки синтезируются в виде предшественников - прокаспаз, активируемых протеолитическим расщеплением, суть которого состоит в отсоединении N-концевого домена и разделении оставшейся молекулы на две части, в последствии ассоциирующиеся в димеры и тетрамеры.
Инициаторные каспазы необходимы для активации эффекторной группы, которая проявляет протеолитическую активность в отношении различных жизненно важных клеточных белков. К субстратам каспаз второго эшелона относятся:
- ферменты репарации ДНК;
- игибитор белка р-53;
- поли-(ADP-рибозо)-полимераза;
- ингибитор ДНК-азы DFF (разрушение этого белка приводит к активации эндонуклеазы CAD) и др.
Общее количество мишеней эффекторных каспаз насчитывает более 60 белков.
Ингибирование апоптоза клеток еще возможно на стадии активации инициаторных прокаспаз. Когда эффекторные каспазы вступают в действие, процесс становится необратимым.
Пути активации апоптоза
Передача сигнала для запуска апоптоза клетки может быть осуществлена двумя путями: рецепторным (или внешним) и митохондриальным. В первом случае процесс активируется через специфические рецепторы смерти, воспринимающие внешние сигналы, которыми служат белки семейства TNF или Fas-лиганды, расположенные на поверхности Т-киллеров.
В состав рецептора входит 2 функциональных домена: трансмембранный (предназначенный для связи с лигандом) и ориентированный внутрь клетки "домен смерти", индуцирующий апоптоз. Механизм рецепторного пути основывается на образовании DISC-комплекса, активирующего инициаторные каспазы 8 или 10.
Сборка начинается со взаимодействия домена смерти с внутриклеточными адапторными белками, которые, в свою очередь, связывают инициаторные прокаспазы. В составе комплекса последние превращаются в функционально-активные каспазы и запускают дальнейший апоптозный каскад.
Механизм внутреннего пути основан на активации протеолитического каскада особыми митохондриальными белками, выброс которых контролируется внутриклеточными сигналами. Выход компонентов органоидов осуществляется через образование огромных пор.
Особая роль в запуске принадлежит цитохрому с. Попадая в цитоплазму, этот компонент электротранспортной цепи связывается с белком Apaf1 (апоптотический фактор активации протеаз), что приводит к активации последнего. Затем Apaf1 связывают инициаторные прокаспазы 9, которые по механизму каскада запускают апоптоз.
Контроль внутреннего пути осуществляется особой группой белков семейства Bcl12, которые регулируют выход межмембранных компонентов митохондрий в цитоплазму. В составе семейства имеются как проапоптические, так и антиапоптические белки, баланс между которыми и определяет, будет ли запущен процесс.
К одним из мощных факторов, запускающих апоптоз по митохондриальному механизму, относятся реактивные формы кислорода. Еще одним значимым индуктором является белок р53, который активирует митохондриальный путь при наличии ДНК-повреждений.
Иногда запуск апоптоза клеток сочетает в себе сразу два пути: как внешний, так и внутренний. Последний обычно служит для усиления рецепторной активации.
Инициация апоптоза может происходить посредством внешних или внутриклеточных факторов. Например, в результате гипоксии, гипероксии, субнекротического поражения химическими или физическими агентами, перекрёстного связывания соответствующих рецепторов, нарушения сигналов клеточного цикла, удаления факторов роста и метаболизма и т. д. Несмотря на разнообразие инициирующих факторов, выделяются два основных пути трансдукции сигнала апоптоза: рецептор-зависимый сигнальный путь с участием рецепторов гибели клетки и митохондриальный путь.
Рецептор-зависимый сигнальный путь
Схема передачи сигналов апоптоза при посредстве рецепторов смерти CD95, TNFR1 и DR3
Процесс апоптоза часто начинается с взаимодействия специфических внеклеточных лигандов с рецепторами клеточной гибели, экспрессированными на поверхности клеточной мембраны. Рецепторы, воспринимающие сигнал апоптоза, относятся к суперсемейству TNF-рецепторов. Наиболее изученными рецепторами смерти, для которых описана и определена роль в апоптозе, являются CD95 и TNFR1. К дополнительным относятся CARI, DR3, DR4 и DR5.
Все рецепторы смерти представляют собой трансмембранные белки, характеризующиеся наличием общей последовательности из 80 аминокислот в цитоплазматическом домене. Данная последовательность называется доменом смерти и является необходимой для трансдукции сигнала апоптоза. Внеклеточные участки рецепторов смерти взаимодействуют с тримерами лигандов. Тримеры лигандов в результате взаимодействия тримеризуют рецепторы смерти. Активированный таким образом рецептор взаимодействует с соответствующим внутриклеточным адаптером. Для рецептора CD95 адаптером является FADD. Для рецепторов TNFR1 и DR3 адаптером является TRADD.
Адаптер, ассоциированный с рецептором смерти, вступает во взаимодействие с эффекторами пока ещё неактивными предшественниками протеаз из семейства инициирующих каспаз с прокаспазами. В результате цепочки взаимодействия «лиганд-рецептор-адаптер-эффектор» формируются агрегаты, в которых происходит активация каспаз. Данные агрегаты именуются апоптосомами, апоптозными шаперонами или сигнальными комплексами индуцирующими смерть. Примером апоптосомы может служить комплекс FasL-Fas-FADD-прокаспаза-8, в котором активируется каспаза-8.
Рецепторы смерти, адаптеры и эффекторы взаимодействуют между собой сходными по структуре доменами: DD, DED, CARD. DD участвует во взаимодействии рецептора Fas с адаптером FADD и во взаимодействии рецепторов TNFR1 или DR3 с адаптером TRADD. Посредством домена DED осуществляется взаимодействие адаптера FADD с прокаспазами −8 и −10. Домен CARD участвует во взаимодействии адаптера RAIDD с прокаспазой-2.
Посредством рецепторов смерти могут быть активированы три инициирующие каспазы: −2; −8 и −10. Активированные инициирующие каспазы далее участвуют в активации эффекторных каспаз.
Митохондриальный сигнальный путь
Митохондриальный сигнальный путь апоптоза реализуется в результате выхода апоптогенных белков из межмембранного пространства митохондрий в цитоплазму клетки. Высвобождение апоптогенных белков, предположительно, может осуществляться двумя путями: за счёт разрыва митохондриальной мембраны или же путём открытия высокопроницаемых каналов на внешней мембране митохондрий.

Модель образования апоптосомы «Цитохром c Apaf-1 CARD прокаспаза-9». Активированная таким образом каспаза-9 рекрутирует прокаспазу-3, которая в свою очередь активируется до каспазы-3
Разрыв внешней мембраны митохондрий объясняется увеличением объема митохондриального матрикса. Данный процесс связывают с раскрытием пор митохондриальной мембраны, приводящим к снижению мембранного потенциала и высокоамплитудному набуханию митохондрий вследствие осмотического дисбаланса. Поры диаметром 2,6-2,9 нм способны пропускать низкомолекулярные вещества массой до 1,5 кДа. Раскрытие пор стимулируют следующие факторы: неорганический фосфат; каспазы; SH-реагенты; истощение клеток восстановленным глутатионом; образование активных форм кислорода; разобщение окислительного фосфорилирования протонофорными соединениями; увеличение содержания Ca в цитоплазме; воздействие церамида; истощение митохондриального пула АТФ и др.
В качестве альтернативного пути выхода апоптогенных белков из межмембранного пространства митохондрий рассматривается вариант образования белкового канала во внешней митохондриальной мембране. Так или иначе, в цитоплазму высвобождаются: цитохром c белок с молекулярной массой 15 кДа; прокаспазы −2, −3 и −9; AIF флавопротеин с молекулярной массой 57 кДа.
Цитохром c в цитоплазме клетки участвует в формировании апоптосомы вместе с белком Apaf-1. Предварительно, Apaf-1 претерпевает конформационные изменения в результате реакции, протекающей с затратой энергии АТФ. Предполагается, что трансформированный Apaf-1 приобретает способность связывать цитохром c. К тому же открывается доступ CARD-домена Apaf-1 для прокаспазы-9. В итоге происходит олигомеризация не менее 8 субъединиц трансформированного белка Apaf-1 с участием цитохрома c и прокаспазы-9. Так образуется апоптосома, активирующая каспазу-9. Зрелая каспаза-9 связывает и активирует прокаспазу-3 с образованием эффекторной каспазы-3. Высвобождающийся из межмембранного пространства митохондрий флавопротеин AIF является эффектором апоптоза, действующим независимо от каспаз.
Другие пути индукции апоптоза
Стоит отметить, что реализация апоптоза может происходить в результате комбинированного действия двух основных сигнальных путей рецептор-зависимого и митохондриального. Помимо этого, существует ряд менее распространённых механизмов инициации апоптоза. Например, за счёт активации прокаспазы-12, локализованной в эндоплазматическом ретикулуме. Высвобождение и активация прокаспазы-12 при этом обусловлены нарушениями внутриклеточного гомеостаза ионов кальция. Активация апоптоза также может быть связана с нарушением адгезии клеток.
В качестве ещё одного фактора индукции апоптоза рассматривается атака инфицированных клеток цитотоксическими Т-лимфоцитами, которые, помимо активации Fas-рецептора, способны секретировать перфорин вблизи мембраны заражённой клетки. Перфорин, полимеризуясь, образует трансмембранные каналы, через которые внутрь клетки поступают лимфотоксин-альфа и смесь сериновых протеаз. Далее гранзим B активирует каспазу-3 и запускается каспазный каскад.
Возможна инициация клеточной смерти при высвобождении лизосомальных протеаз катепсинов. К примеру, каспаза-8 вызывает выход из лизосом активного катепсина B, который затем расщепляет регуляторный белок Bid. В результате образуется активный белок t-Bid, активирующий в свою очередь проапоптозный белок Bax.

Общая схема «классического» апоптоза млекопитающих
Эффекторная фаза
В течение эффекторной фазы различные инициирующие пути конвертируются в один общий путь апоптоза. Как правило, происходит активация каскада белков-эффекторов и регулирующих их белков-модуляторов. Основными эффекторами апоптоза являются каспазы. В процессе активации они запускают каспазный каскад: сложно переплетённые цепочки взаимодействий инициирующих и эффекторных каспаз.
Каспазный каскад
Каспазы представляют собой цистеиновые протеазы, которые расщепляют аминокислотные последовательности после остатка аспарагиновой кислоты. Каспазы образуются за счёт активации прокаспаз, в составе которых выделяют 3 домена: регуляторный N-концевой домен, большую и малую субъединицы. Активация происходит путём протеолитического процессинга: все три домена расщепляются, отделяется продомен, а оставшиеся большая и малая субъединицы ассоциируются, образуя гетеродимер. Два гетеродимера в дальнейшем формируют тетрамер полноценную каспазу с двумя каталитическими участками.
Каспазы обнаружены во большинстве живых организмов. У млекопитающих идентифицировано 13 каспаз. Часть из них в апоптозе не участвует. Остальные каспазы, которые участвуют в апоптозе, разделяют на инициаторные и эффекторные. Инициаторные каспазы активируют эффекторные каспазы, которые в свою очередь провоцируют и непосредственно участвуют в трансформации клетки. В итоге морфологические и биохимические изменения приводят к гибели клетки по типу апоптоза.
Одна из основных функций эффекторных каспаз заключается в прямом и опосредованном разрушении клеточных структур. Гидролизу подвергаются белки ядерной ламины, разрушается цитоскелет, расщепляются белки, регулирующие клеточную адгезию. Другой важной функцией эффекторных каспаз является инактивация белков, блокирующих апоптоз. В частности расщепляется ингибитор DFF, препятствующий активации апоптозной ДНКазы CAD. Разрушению подвергаются и антиапоптозные белки семейства Bcl-2. Наконец, в результате действия эффекторных каспаз происходит диссоциация регуляторных и эффекторных доменов, участвующих в репарации ДНК, мРНК-сплайсинга и ДНК-репликации.
Дополнительные эффекторы апоптоза
Помимо каспаз существуют и другие эффекторы апоптоза. Например, флавопротеин AIF, высвобождающийся из межмембранного пространства митохондрий, действует по независимому от каспаз пути. Попадая в клеточное ядро, AIF вызывает конденсацию хроматина и активирует эндонуклеазы, которые участвуют в фрагментации ДНК. На основании экспериментальных данных установлено, что апоптоз, протекающий в присутствии AIF, не предотвращается ингибитором каспаз. В качестве эффекторов апоптоза также рассматриваются кальпаины представители семейства цитозольных Ca-активируемых цистеиновых протеаз. Их роль в апоптозе пока слабо охарактеризована.
Деградационная фаза
Итогом программируемой клеточной гибели вне зависимости от изначального инициирующего воздействия является деградация клетки путём фрагментации на отдельные апоптотические тельца, ограниченные плазматической мембраной. Фрагменты погибшей клетки обычно очень быстро фагоцитируются макрофагами либо соседними клетками, минуя развитие воспалительной реакции.
Морфологические изменения
Условно деградацию погибающей клетки можно разделить на три последовательных фазы: высвобождения, блеббинга и конденсации. Деградация большинства клеток начинается с высвобождения прикреплений внеклеточного матрикса и реорганизации фокальной адгезии. Внутри погибающей клетки деполимеризуются микротрубочки цитоскелета. Внутриклеточные актиновые микрофиламенты реорганизуются в связанные с мембраной периферийные кольцевые пучки. В итоге клетка приобретает округлую форму. Следующая за высвобождением, стадия блеббинга, характеризуется сокращением периферийных актиновых колец. В результате сокращений клеточная мембрана образует вздутия, клетка как бы «кипит». Процесс блеббинга энергозависим и требует большого количества АТФ. Фаза блеббинга в нормальных условиях завершается примерно через час. В итоге клетка фрагментируется на маленькие апоптотические тела, либо целиком конденсируется, округляясь и уменьшаясь в размерах.
Биохимические изменения
На молекулярном уровне одним из последствий апоптоза является фрагментация ДНК с участием нуклеаз. Изначально образуются крупные фрагменты с 30 000-700 000 пар оснований, которые в дальнейшем расщепляются в межнуклеосомной области на отрезки по 180-190 пар оснований или кратные этим величинам. Фрагментация ДНК является характерным, но не обязательным признаком апоптоза, так как существуют наблюдения, в ходе которых процесс фрагментации ядра протекал без сопутствующей фрагментации ДНК.
Ещё одним существенным последствием апоптоза является экспрессия на внешней стороне плазматической мембраны специфических молекулярных маркеров, распознаваемых фагоцитирующими клетками: тромбоспондина; фосфатидилсерина и других фосфолипидов, содержащих фосфосерин.